Introduction
Pour beaucoup de personnes et même pour un certain nombre de thérapeutes/coachs, la perte de poids ne serait qu’une question de balance énergétique : si je mange plus que mes besoins, je grossis. Inversement, si j’apporte moins d’énergie à mon corps que ce qu’il en dépense au quotidien, je perds du poids. Cette affirmation est basée sur un modèle mécanistique et sur la 1ère loi de la thermodynamique qui reprend la formule de Lavoisier pour affirmer que « l’énergie ne peut ni être créée ni détruite, mais seulement transformée d’une forme à une autre ». Autrement dit, l’énergie totale d’un système isolé reste constante au fil du temps.
Mais c’est précisément la que la limite intervient, le corps humain étant loin d’être un système isolé : de nombreux facteurs peuvent ainsi influencer la dépense énergétique et le métabolisme de base. Parmi ces facteurs, on retrouve notamment l’activité thyroïdienne, la thermogénèse adaptative ou encore le métabolisme mitochondrial. Les équations permettant d’estimer le métabolisme de base et la dépense énergétique quotidienne d’un individu peuvent donc être très approximatives en pratique. La seule méthode de mesure fiable en pratique restant à ce jour la calorimétrie indirecte.
Deux personnes du même gabarit et ingérant la même quantité d’énergie peuvent donc observer des variations de poids tout à fait différentes.
Nous allons voir dans cet article que le microbiote intestinal pourrait un jouer un rôle central en ce qui concerne la capacité du corps à utiliser une quantité plus ou moins importante de l’énergie que nous lui apportons via l’alimentation.
Le microbiote étant un sujet nouveau et encore relativement mal compris, je tenterais de rester prudent dans mes écrits. En tant que lecteur, je vous invite également, encore plus que d’habitude compte tenu du sujet, à prendre mes propos avec des pincettes et avec du recul.
Ceci étant dit, nous voila partis pour une plongée dans le monde fascinant des micro-organismes et de notre système digestif 😊
Le microbiote intestinal, c’est quoi exactement ?
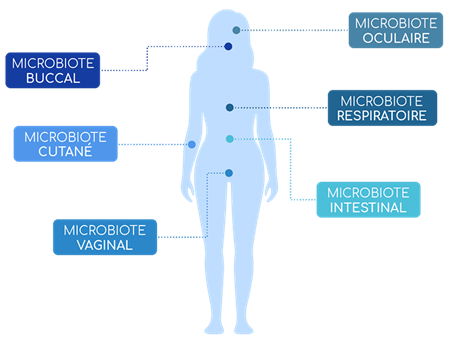
Un microbiote, au sens large du terme, désigne l’ensemble des micro-organismes – bactéries, virus, parasites et champignons – vivant dans un environnement spécifique. Au sein du corps humain, on retrouve une multitude de microbiotes, notamment au niveau des muqueuses : microbiote digestif, pulmonaire, vaginal ou encore buccal. Le microbiote intestinal est sans aucun doute possible le plus étudié des microbiotes. Il est aussi celui qui contient, et de loin, le plus de bactéries au sein de notre organisme. Les chercheurs estiment en effet que l’on trouve environ un millier d’espèces différentes de bactéries au sein de notre organisme, pour un nombre total de bactéries d’environ 40 000 milliards. En d’autres termes, nous possédons à peu près autant de bactéries dans notre corps que de cellules (1). Le microbiote humain comporterait également 3 à 4 millions de gènes, contre « seulement » 20 à 25 000 pour le génome humain (2). Son impact sur notre santé est donc absolument crucial.
Ce microbiote intestinal est principalement situé au niveau du colon (3) ou l’activité microbienne est bien plus riche et plus dense qu’au niveau de l’intestin grêle. On estime le nombre de bactéries à 103 dans le duodénum, 104 dans le jéjunum, 107 dans l’iléon et enfin 1012 dans le colon (4).
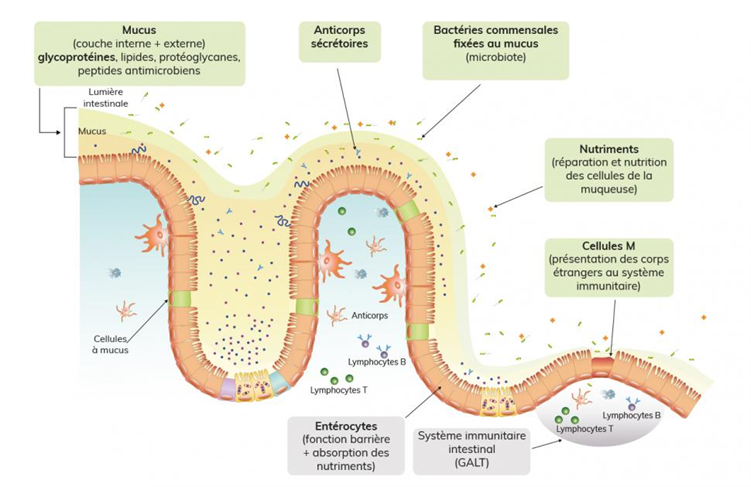
D’un point de vue structurel, le microbiote n’est pas en contact direct avec les cellules de l’intestin grêle (entérocytes) et du colon (colonocytes). Le microbiote est situé en partie dans la lumière intestinale du tube digestif et en partie au contact du mucus. Le mucus représente une forme de gel composé à 95% d’eau ainsi que d’électrolytes, de lipides et de protéines spécifiques produites par les cellules de Gobelet que l’on appelle des mucines (5). Dans le colon, ce mucus est composé d’une couche interne collée à la membrane intestinale (cf. schéma ci-dessous) et normalement stérile ainsi que d’une couche externe qui va être colonisée par certaines bactéries. Le mucus permet donc d’assurer l’intégrité et la stérilité de la membrane intestinale. De plus, il semblerait que les propriétés de ce mucus soient directement impactées par la composition du microbiote, comme semblent le démontrer des travaux effectués sur des souris identiques au niveau génétique mais dont la composition du microbiote avait été modifiée par des environnements différents (6).
Comment le microbiote influence-t-il l’extraction calorique ?
Cela permet d’évoquer un point fondamental en ce qui concerne le microbiote intestinal : bien qu’il reste globalement stable, il peut malgré tout sensiblement évoluer à l’âge adulte et ce en fonction de nombreux paramètres environnementaux comme l’alimentation, le sommeil, la pollution ou encore le stress chronique. De plus, le microbiote intestinal est spécifique à chaque individu de sorte qu’il constitue l’équivalent d’une « empreinte microbienne ».
Dès lors, nous en revenons donc à la question centrale de cet article : deux individus avec un microbiote différents peuvent ils utiliser différemment l’énergie théoriquement apportée par les aliments consommés ?
Les premières études ayant suggéré le rôle du microbiote en ce qui concerne l’extraction calorique des aliments remontent au début de 21ème siècle et furent menées par l’équipe du docteur Jeffrey Gordon de l’université de médecine de Washington.
Ils parvinrent notamment à montrer, dès 2006 (7), que la transplantation d’un certain microbiote chez des axéniques (dépourvues de microbiote) modifiées génétiquement pouvait considérablement impacter la composition corporelle. Pour y parvenir, ils ont ainsi greffé un microbiote de type « obèse » à 5 souris axénique et un microbiote dit « sain » à 4 autres souris. Sans que ces différents microbiotes impactent la quantité de nourriture consommée par les souris qui est restée équivalente entre les 2 groupes, les chercheurs ont constaté une augmentation bien plus importante du taux de masse grasse chez les souris ayant recu le microbiote de type « obèse ». Bien que cette étude ait été effectuée sur des modèles animaux et avec un très faible nombre de sujets, les chercheurs concluent en disant que « la modification de l’efficacité de l’assimilation énergétique provenant de l’alimentation, induite par des changements dans l’écologie microbienne intestinale, n’a pas besoin d’être importante pour contribuer à l’obésité, étant donné que de légers changements dans l’équilibre énergétique, sur une année, peuvent entraîner des variations significatives du poids corporel ».
Et il semblerait que cette tendance ait pu être confirmée dans d’autres études. L’une d’entre elles, publiée en 2023 (8), a permis de regrouper 17 participants au sein d’une étude randomisée en crossover, c’est-à-dire ou chaque participant est son propre contrôle. Il était demandé aux participants de consommer pendant 10 jours soit une diète de type occidentale soit une diète favorable à la composition du microbiote, c’est-à-dire riche en fibres et en aliments peu transformés (fruits, noix, légumes etc..). Une fois les 10 jours d’intervention passés, une période dite « de wash-out » était observée pendant 14 jours puis les participants adoptaient l’autre diète, la encore pendant 10 jours. Alors que la plupart des paramètres environnementaux (sommeil, apports caloriques, dépenses etc..) étaient contrôlés et gardés constants, l’alimentation riche en fibres a conduit à une réduction d’énergie assimilée d’en moyenne 116 ± 56 kcal (P < 0.0001). Autrement dit, une alimentation de mauvaise qualité pourrait représenter un surplus de 116kcal réellement assimilées par le corps par rapport à une alimentation de bonne qualité apportant la même quantité d’énergie théorique.
Ces observations avaient déjà été faites en 1997 au sein d’une étude randomisée la aussi menée sur 17 sujets (9). Au sein de cette étude, la proportion de fibres dans l’alimentation était inversement corrélée à l’énergie métabolisable. Ainsi, au plus l’alimentation était riche en fibres, au moins le corps était capable d’utiliser l’énergie issue de l’alimentation, en partie grâce à une digestibilité réduite des graisses et des protéines. En 2011, une autre étude (10) a permis de mettre en évidence qu’une modification de l’apport calorique de 2400 à 3400kcal était susceptible d’engendrer des modifications majeures au niveau des populations bactériennes avec notamment une modification du ratio firmicutes/Bacteroidetes. Cette modification était associée dans l’étude à une augmentation de l’extraction calorique d’environ 150kcal/jour. En parlant du ratio firmicutes/Bacteroidetes, celui-ci s’avère bien souvent accru dans le cadre de l’obésité (11)(12), bien que certaines études ne retrouvent pas de modifications à ce niveau (13). De même, il se pourrait que l’obésité soit associée à une réduction de la diversité microbienne (14), bien que cela ne soit pas retrouvé dans toutes les études (15).
Les acides gras à chaîne courte (AGCC) : le chaînon manquant
Mais alors comment expliquer que le microbiote puisse potentiellement jouer un tel rôle dans l’extraction des calories issues des aliments ?
Pour comprendre cela, il faut s’attarder un petit moment sur des molécules bien particulières que l’on appelle les acides gras à chaîne courte (AGCC). Ces AGCC sont produits par la fermentation des fibres alimentaires par le microbiote intestinal et sont au nombre de 3 : le butyrate, le propionate et l’acétate.
D’après les études récentes, ces AGCC pourraient avoir de nombreux rôles et de nombreuses conséquences d’un point de vue métabolique.
En 1er lieu, une méta-analyse (16) de 2023 regroupant 23 études ayant cherché à augmenter les concentrations d’AGCC nous apprend que des concentrations accrues d’AGCC ont permis de significativement réduire les concentrations d’insuline à jeun (SMD = -0.15 ; 95%CI = -0.29 to -0.01, P = 0.04) ainsi que les valeurs de HOMA-IR (P < 0.00001). Cette observation recoupe les résultats de nombreuses études ou un apport conséquent/augmenté en fibres alimentaires est très fréquemment associé à une augmentation de la sensibilité à l’insuline, aussi bien chez des sujets victimes de diabète de type 2 que chez des sujets sains (17)(18). A noter toutefois que ce lien entre fibres alimentaires et sensibilité à l’insuline pourrait être médié par l’indice de masse corporelle (IMC), le tour de taille et le taux de masse grasse.
La plupart des études menées sur le sujet suggèrent également qu’une production accrue d’AGCC pourrait faire baisser les niveaux d’inflammation et améliorer certains paramètres lipidiques (19). De plus, les AGCC pourraient exercer un rôle protecteur sur les cellules épithéliales de l’intestin grêle et du colon en renforçant les jonctions serrées (20). Dès lors, les AGCC pourraient lutter contre l’hyperperméabilité intestinale, celle-ci pouvant être associée au passage dans le sang de molécules pro-inflammatoires nommées lipopolysaccharides (LPS). Ces LPS seraient notamment impliqués dans l’inflammation de bas grade, la résistance à l’insuline et la prise de poids.
En plus de ces paramètres métaboliques, les AGCC pourraient contribuer à réguler l’appétit et la satiété en augmentant la production de certaines hormones comme le GLP-1, la leptine ou le peptide YY. Ces données sont retrouvées de façon systématique chez l’animal, aussi bien la souris que le cochon (21)(22). Globalement, les effets de la supplémentation en AGCC semblent bien plus prometteurs chez les animaux que sur les humains. Chez l’animal, une supplémentation en AGCC permet fréquemment de réduire l’appétit, l’apport calorique et l’inflammation tout en augmentant la dépense calorique, la thermogénèse et la conversion du tissu adipeux blanc en tissu adipeux brun. Tous ces effets contribuent dans de nombreuses études animales à réguler efficacement le poids et à prévenir la prise de poids et les troubles métaboliques (23)(24).
Malheureusement, les données chez l’humain sont bien moins concluantes : il semblerait ainsi que la supplémentation en AGCC ne soit pas en mesure de réguler l’appétit et ainsi de baisser l’apport calorique (25(26). Cela pourrait s’expliquer par le fait que les AGCC consommées par voie orale sont absorbés au niveau de l’intestin grêle et non du colon qui correspond théoriquement à l’endroit où ils sont produits. Une étude randomisée ingénieuse (27) a ainsi réussi à contourner le problème : 60 sujets en situation de surpoids étaient ainsi répartis aléatoirement au sein de 2 groupes et ce pour une durée de 24 semaines. Le groupe test consommait 10gr/jour de propionate spécifiquement conçu pour n’être absorbé qu’au niveau du colon. Le groupe contrôle consommait un placebo à base d’inuline. Les chercheurs ont ainsi pu démontrer que la consommation en aigue de 10gr de propionate permettait d’augmenter les niveaux de GLP-1 et de PYY et ainsi de réduire l’apport calorique du repas de 14% (162kcal). Toutefois, la réponse à moyen-long terme s’est avérée moins importante puisque entrainant seulement une réduction de 73kcal de l’apport calorique moyen au cours d’un repas. La prise de poids était cependant moins importante dans le groupe test qu’au sein du groupe contrôle, soulignant le rôle potentiellement important des AGCC et de la consommation de fibres en ce qui concerne le contrôle du poids corporel et du métabolisme.
Très récemment, une étude randomisée (28) conduite auprès 46 adultes en situation de surpoids et pour la moitié victime de diabète de type 2 semble confirmer les résultats de l’étude précédente. Cette fois, le groupe test consommait 1875mg de butyrate de sodium/jour pendant 12 semaines alors que le groupe témoin recevait un placebo. Alors que tous les participants suivaient une alimentation légèrement hypocalorique, la perte de poids chez les sujets dépourvus de diabète s’est avérée bien plus importante dans le groupe butyrate qu’au sein du groupe contrôle (-7.0 ± 3.0 vs. -3.2 ± 1.6 kg ; p = 0.001). Chez les sujets diabétiques, la consommation de butyrate a permis de faire baisser les valeurs de triglycérides (-0.36 ± 0.47 vs. +0.08 ± 0.30 mmol/L ; p = 0.012) et d’améliorer le contrôle glycémique de façon bien supérieure au groupe placebo.
Enfin, une méta-analyse (29) de 2022 regroupant 1400 participants suggèrent que la supplémentation en fibres solubles permettrait d’atteindre une perte de poids plus importante qu’au sein des groupes contrôles (MD −1.25 kg, 95% CI −2.24, −0.25 ; p = 0.01; I2 = 0%). Ces résultats sont en accord avec ceux d’une autre méta-analyse de 2017 (30) qui démontrait qu’une supplémentation en fibres solubles chez des personnes en situation de surpoids ou d’obésité permettait d’engendrer une perte de poids bien supérieure à ce qui était retrouvé au sein des groupes contrôles (-2.52 kg ; 95% CI : -4.25, -0.79 kg ; P = 0.004).
Au global, il semble de plus en plus certain que le microbiote et que la consommation de fibres joue un rôle important sur la perte de poids en régulant notamment l’extraction calorique, la sensation de faim et certains paramètres métaboliques. Toutefois, nous manquons encore de données solides et d’études long-terme chez l’humain afin de pouvoir tirer des conclusions définitives sur le sujet.
L’axe intestin-cerveau et la régulation du comportement alimentaire
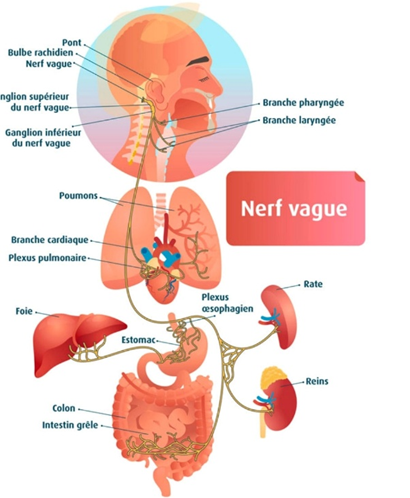
Mais au-delà de la simple consommation de fibres et de son impact direct sur la composition du microbiote, il faut garder en tête que le microbiote communique en permanence avec le cerveau par l’intermédiaire du nerf vague ; les anglais parlent de « gut-brain axis », littéralement l’axe intestin – cerveau. Le rôle de ce nerf vague est majoritairement de transmettre les informations issues des différents organes du corps en direction du cerveau et ce afin de réguler le système nerveux autonome dit parasympathique (en charge du repos et la digestion). Le nerf vague est composé à 80% de fibres afférentes ; c’est-à-dire qui remontent des organes jusqu’au cerveau.
Cela possède un enjeu majeur en ce qui concerne la régulation des envies et des comportements alimentaires : en effet, il est probable que la composition du microbiote puisse directement influencer le comportement alimentaire en favorisant notamment l’appétence pour des produits sucrés. Et pour cause, 95% de la sérotonine étant produite au niveau de l’intestin, il y a fort à parier qu’une perturbation du microbiote intestinal puisse réduire la production de sérotonine et donc engendrer des troubles de l’humeur et du comportement alimentaire, avec notamment une appétence plus importance pour les produits sucrés comme le chocolat, les biscuits ou les gâteaux industriels. Il convient toutefois de garder en tête que l’appétence pour le sucré peut provenir d’énormément de facteurs (stress, privation de sommeil, constructions mentales) pouvant interagir les uns avec les autres.
En parlant de troubles de l’humeur, rappelons également que le microbiote semble jouer un rôle important dans la survenue de certains troubles de l’humeur ou d’épisodes de dépression. Les bactéries pourraient ainsi communiquer avec le cerveau par l’intermédiaire du nerf vague et ainsi moduler la production de neurotransmetteurs fondamentaux pour la régulation de l’humeur comme la sérotonine, le glutamate ou le GABA. La dysbiose intestinale est ainsi très souvent retrouvée chez des personnes en situation de dépression (31)(32). La dysbiose intestinale pourrait, comme nous l’avons vu précédemment, engendrer un état d’inflammation en réduisant la production d’AGCC et en favorisant à l’inverse le passage dans le sang de molécules pro-inflammatoires comme les LPS. Or l’inflammation peut réduire la production de sérotonine en détournant le tryptophane vers une voie alternative appelée kynurénine. L’inflammation de bas grade et la neuro-inflammation sont ainsi souvent retrouvées chez des sujets victimes de dépressions plus ou moins importantes (33).
De plus, il a été mis en évidence sur des modèles animaux qu’une greffe de microbiote issu d’un animal souffrant de traits dépressifs pouvait, chez un animal initialement en bonne santé, engendrer à son tour le développement de comportements dépressifs (34).
Il convient donc de ne pas négliger le rôle du microbiote en ce qui concerne les troubles de l’humeur/psychiques et donc les conséquences potentielles sur les apports et les comportements alimentaires. En effet, de nombreux travaux démontrent que la dépression et les troubles de l’humeur peuvent significativement augmenter les apports caloriques et par conséquent le poids corporel par des mécanismes de « réconfort émotionnel » (35)(36).
6 conseils de bon sens pour bichonner son microbiote
Bien que le microbiote intestinal regorge encore de mystères et de secrets, il ne fait désormais plus aucun doute qu’il est en capacité d’influencer le contrôle du poids et les différents paramètres métaboliques associés et ce par l’intermédiaire de nombreux processus physiologiques.
Puisque nous avons au chapitre sur les AGCC que la supplémentation directe ne semblait pas vraiment prometteuse chez l’humain, la question à un million d’euros demeure la suivante : que faire pour optimiser son microbiote et ainsi mettre toutes les chances de son côté pour optimiser sa santé et ne pas prendre de poids ?
La encore, nous n’avons probablement pas encore toutes les réponses à ce jour. Cependant, de nombreuses recommandations validées scientifiquement recoupent des pratiques de bon sens en matière de santé. Voici les principaux facteurs pouvant influer sur la composition du microbiote :
- La consommation de fibres : en tant que nourriture directe du microbiote intestinal, les fibres alimentaires s’avèrent absolument cruciales pour disposer d’un bon écosystème intestinal. Une consommation accrue et suffisante de fibres est systématiquement associée dans les études à un risque réduit de mortalité toutes causes confondues, de maladies métaboliques et de troubles cardio-vasculaires. La consommation journalière recommandée est d’environ 25-30gr, bien que ce chiffre puisse varier en fonction des pays (37). Il convient également de varier au maximum les végétaux consommés et ce afin de pouvoir disposer de sources de fibres différentes ; celles-ci se distinguent en effet entre fibres solubles (pectines, béta-glucanes, inuline, gommes : présentes notamment dans les fruits, les légumes et l’avoine) et les fibres insolubles (cellulose, amidon résistant, lignine : retrouvées notamment dans les céréales complètes et les légumineuses). Le fait de varier au maximum votre consommation de fruits, de légumes, de céréales, d’épices et d’aromates vous permettra également de maximiser votre apport en antioxydants, vitamines, minéraux et polyphénols.
- Eviter les aliments ultra-transformés : En parallèle d’une consommation importante de fibres, un microbiote de qualité passe également par une faible consommation d’aliments ultra-transformés : ceux-ci sont en effet riches en graisses saturées voir trans, en sucres raffinés et en additifs en plus d’être pauvres en fibres. Parmi ces produits, on retrouve par exemple les gâteaux industriels, les bonbons, les plats préparés (pasta box, noodles etc..) ou encore les charcuteries. Une consommation importante de ces produits est associée à une augmentation des risques de dysbiose intestinale (38)(39) ainsi qu’à des niveaux d’inflammation dans l’organisme accrus. Mais à ce niveau, je ne pense pas apprendre grand-chose à qui que ce soit… 😉
- Pratiquer une activité physique régulière : La encore, un conseil de bon sens qui semble démontré par les études scientifiques. Dans ces dernières, la pratique d’une activité physique régulière et d’intensité modérée permettrait en effet d’augmenter la diversité microbienne par rapport à des personnes sédentaires (40). La pratique d’une activité physique régulière favoriserait également la production d’AGCC par les bactéries et serait ainsi garante de l’intégrité de la muqueuse intestinale, réduisant ainsi les risques d’inflammation de bas grade (41). Pour ce qui est modalités, il se pourrait que la pratique d’une activité physique en endurance soit plus pertinente que l’entrainement en force, bien que plus de données soient nécessaires pour tirer de réelles conclusions sur le sujet. Ce qui est certain cependant, c’est que l’entrainement en endurance à haute intensité s’avère bien souvent délétère pour le microbiote et pour la sphère digestive en étant à l’origine de dommages importants au niveau de la muqueuse intestinale (42).
- Eviter le stress chronique : Trop souvent sous-estimé lorsqu’il s’agit de la santé intestinale et digestive, le stress chronique et l’aspect émotionnel s’avèrent pourtant très importants. En effet, le stress chronique ainsi qu’un épisode de vie très stressant comme a pu l’être la crise COVID chez les soigneurs sont tout deux associés à des modifications durables du microbiote (43)(44). Chez la souris, le stress psychosocial chronique est associé à une altération de la muqueuse intestinale et du mucus (45). Il existe qui plus est un véritable cercle vicieux entre microbiote et stress chronique puisqu’une altération du microbiote pourrait également contribuer à l’apparition de troubles anxieux (46). Il est cependant trop tôt pour affirmer avec certitude que des pratiques comme la méditation ou la cohérence cardiaque permettent de moduler positivement le microbiote. Toutefois, une étude effectuée chez des individus ayant participé à un stage de méditation de 9 jours (47) a constaté une augmentation de certaines bactéries bénéfiques de type Prevotella ainsi qu’une diminution de la production de cytokines pro-inflammatoires et du stress perçu par les participants. Chez des moines tibétains pratiquant la méditation de longue date, une richesse plus importante du microbiote a pu être constatée par rapport à leurs homologues tibétains ne pratiquant pas la méditation (48). Ces différences étaient associées à un moindre risque de troubles anxieux et dépressifs ainsi que de maladies cardio-vasculaires.
- Dormir suffisament : De nombreux français ne dorment malheureusement pas suffisament au quotidien. Les effets du manque chronique de sommeil sur les troubles métaboliques et cardio-vasculaires sont depuis longtemps démontrés. Il se pourrait que ces effets soient médiés par le microbiote intestinal ; en effet, nous savons désormais que les bactéries du microbiote sont sensibles aux perturbations du rythme biologique et au décalage horaire (49). Chez l’animal, la dette de sommeil est associée à une dysbiose intestinale ainsi qu’à une augmentation des niveaux d’inflammation et à une augmentation des troubles cognitifs et anxieux (50)(51). Un sommeil insuffisant pourrait ainsi perturber certaines voies de signalisation comme la voie NLRP3 inflammasome et la voie NF-κB (52) et ainsi induire de nombreux effets délétères sur la santé humaine.
- Limiter la consommation de médicaments, d’alcool et de tabac : Tout d’abord, la consommation de tabac est bien entendu associée à une réduction de la diversité microbienne (53) ainsi qu’à une modification de certaines espèces. On retrouverait ainsi chez les fumeurs et les anciens fumeurs une quantité plus importante et anormale de protéobactéries (54), des bactéries à Gram négatif entrainant la production de LPS et qui pourraient, lorsque présentes en excès dans le microbiote, être associées à des troubles inflammatoires et/ou métaboliques. La consommation importante et fréquente d’alcool semble également associée à une réduction de la diversité bactérienne (55) et à une augmentation de la perméabilité intestinale et de l’inflammation de bas grade. Chez le rat, une consommation modérée d’alcool semble également associée à une réduction de la diversité microbienne (56), bien que les études chez l’humain soient plus contrastées. Dans le cas de la bière, une consommation modérée/occasionnelle pourrait même être associée à des bienfaits sur le microbiote (57)(58). Malgré ces impacts potentiellement positifs, une autre étude (59) mentionne que la plus petite consommation possible d’alcool demeure toutefois la plus sure en matière de santé. Enfin, il faut savoir que les antibiotiques, notamment à spectre large, affectent grandement la structure du microbiote puisqu’ils pourraient réduire jusqu’à 30% la diversité microbienne (60). Les inhibiteurs de la pompe à protons (61) pourraient également réduire la diversité microbienne et ainsi augmenter le risque d’infection à clostridium difficile.
Conclusion
Au global, cet article permet de mettre en lumière le rôle significatif que joue probablement le microbiote sur la régulation du métabolisme énergétique et sur la gestion du poids en modulant des processus fondamentaux comme l’extraction calorique, la régulation de l’appétit ou encore l’inflammation.
Pour autant, gardons en tête que la régulation du poids chez une personne est liée à de nombreux facteurs autres que le microbiote. De plus, mon propos ne consiste pas à faire du microbiote l’acteur unique/central de la prise de poids, mais plutôt de mettre en avant le fait que prendre ou perdre du poids n’est pas seulement une question de « calories in vs calories out » ou de volonté.
Malgré le fait que le sujet reste encore relativement récent et mal compris au niveau scientifique, il semblerait qu’un microbiote de qualité soit donc un élément central voire fondamentale d’une perte de poids durable et saine. A cet égard, oubliez les régimes spécifiques pour le microbiote et les compléments alimentaire miracles. Misez plutôt sur du bon sens et sur des éléments validés scientifiquement comme une consommation importante et diversifiée de fibres, un sommeil suffisant, une bonne gestion du stress au quotidien ou encore une activité physique régulière.
Les futures recherches scientifiques permettront peut-être de trouver des outils thérapeutiques plus spécifiques en ce qui concerne la perte de poids et le microbiote mais, pour l’instant, il semble une fois de plus nécessaire de revenir aux fondamentaux et aux piliers d’une bonne santé !
Références scientifiques
- Qin, J., Li, R., Raes, J., Arumugam, M., Burgdorf, K., Manichanh, C., Nielsen, T., Pons, N., Levenez, F., Yamada, T., Mende, D., Li, J., Xu, J., Li, S., Li, D., Cao, J., Wang, B., Liang, H., Zheng, H., Xie, Y., Tap, J., Lepage, P., Bertalan, M., Batto, J., Hansen, T., Paslier, D., Linneberg, A., Nielsen, H., Pelletier, É., Renault, P., Sicheritz-Pontén, T., Turner, K., Zhu, H., Yu, C., Li, S., Jian, M., Zhou, Y., Li, Y., Zhang, X., Li, S., Qin, N., Yang, H., Wang, J., Brunak, S., Doré, J., Guarner, F., Kristiansen, K., Pedersen, O., Parkhill, J., Weissenbach, J., Bork, P., Ehrlich, S., & Wang, J. (2010). A human gut microbial gene catalogue established by metagenomic sequencing. Nature, 464, 59-65. https://doi.org/10.1038/nature08821.
- Sender, R., Fuchs, S., & Milo, R. (2016). Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biology, 14. https://doi.org/10.1371/journal.pbio.1002533.
- Jensen, B., Heyndrickx, M., Jonkers, D., Mackie, A., Millet, S., Naghibi, M., Pærregaard, S., Pot, B., Saulnier, D., Sina, C., Sterkman, L., Van Den Abbeele, P., Venlet, N., Zoetendal, E., & Ouwehand, A. (2023). Small intestine vs. colon ecology and physiology: Why it matters in probiotic administration. Cell Reports Medicine, 4. https://doi.org/10.1016/j.xcrm.2023.101190.
- Sommer, F., & Bäckhed, F. (2013). The gut microbiota–masters of host development and physiology. Nature reviews. Microbiology, 11(4), 227–238. https://doi.org/10.1038/nrmicro2974
- Paone, P., & Cani, P. (2020). Mucus barrier, mucins and gut microbiota: the expected slimy partners?. Gut, 69, 2232 – 2243. https://doi.org/10.1136/gutjnl-2020-322260.
- Jakobsson, H. E., Rodríguez-Piñeiro, A. M., Schütte, A., Ermund, A., Boysen, P., Bemark, M., Sommer, F., Bäckhed, F., Hansson, G. C., & Johansson, M. E. (2015). The composition of the gut microbiota shapes the colon mucus barrier. EMBO reports, 16(2), 164–177. https://doi.org/10.15252/embr.201439263
- Turnbaugh, P. J., Ley, R. E., Mahowald, M. A., Magrini, V., Mardis, E. R., & Gordon, J. I. (2006). An obesity-associated gut microbiome with increased capacity for energy harvest. Nature, 444(7122), 1027–1031. https://doi.org/10.1038/nature05414
- Corbin, K. D., Carnero, E. A., Dirks, B., Igudesman, D., Yi, F., Marcus, A., Davis, T. L., Pratley, R. E., Rittmann, B. E., Krajmalnik-Brown, R., & Smith, S. R. (2023). Host-diet-gut microbiome interactions influence human energy balance: a randomized clinical trial. Nature communications, 14(1), 3161. https://doi.org/10.1038/s41467-023-38778-x
- Baer, D. J., Rumpler, W. V., Miles, C. W., & Fahey, G. C., Jr (1997). Dietary fiber decreases the metabolizable energy content and nutrient digestibility of mixed diets fed to humans. The Journal of nutrition, 127(4), 579–586. https://doi.org/10.1093/jn/127.4.579
- Jumpertz, R., Le, D. S., Turnbaugh, P. J., Trinidad, C., Bogardus, C., Gordon, J. I., & Krakoff, J. (2011). Energy-balance studies reveal associations between gut microbes, caloric load, and nutrient absorption in humans. The American journal of clinical nutrition, 94(1), 58–65. https://doi.org/10.3945/ajcn.110.010132
- Koliada, A., Syzenko, G., Moseiko, V., Budovska, L., Puchkov, K., Perederiy, V., Gavalko, Y., Dorofeyev, A., Romanenko, M., Tkach, S., Sineok, L., Lushchak, O., & Vaiserman, A. (2017). Association between body mass index and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population. BMC Microbiology, 17. https://doi.org/10.1186/s12866-017-1027-1.
- Crovesy, L., Masterson, D., & Rosado, E. (2020). Profile of the gut microbiota of adults with obesity: a systematic review. European Journal of Clinical Nutrition, 1-12. https://doi.org/10.1038/s41430-020-0607-6.
- Duncan, S., Lobley, G., Holtrop, G., Ince, J., Johnstone, A., Louis, P., & Flint, H. (2008). Human colonic microbiota associated with diet, obesity and weight loss. International Journal of Obesity, 32, 1720-1724. https://doi.org/10.1038/ijo.2008.155.
- Chanda, D., & De, D. (2022). Meta-analysis reveals obesity associated gut microbial alteration patterns and reproducible contributors of functional shift. Gut Microbes, 16. https://doi.org/10.1080/19490976.2024.2304900.
- Pinart, M., Dötsch, A., Schlicht, K., Laudes, M., Bouwman, J., Forslund, S., Pischon, T., & Nimptsch, K. (2021). Gut Microbiome Composition in Obese and Non-Obese Persons: A Systematic Review and Meta-Analysis. Nutrients, 14. https://doi.org/10.3390/nu14010012.
- Pham, N., Joglekar, M., Wong, W., Nassif, N., Simpson, A., & Hardikar, A. (2023). Short-chain fatty acids and insulin sensitivity: a systematic review and meta-analysis. Nutrition Reviews, 82, 193 – 209. https://doi.org/10.1093/nutrit/nuad042.de
- Ylönen, K., Saloranta, C., Kronberg-Kippilä, C., Groop, L., Aro, A., & Virtanen, S. (2003). Associations of dietary fiber with glucose metabolism in nondiabetic relatives of subjects with type 2 diabetes: the Botnia Dietary Study.. Diabetes care, 26 7, 1979-85 . https://doi.org/10.2337/diacare.26.7.1979.
- Breneman, C., & Tucker, L. (2012). Dietary fibre consumption and insulin resistance – the role of body fat and physical activity. British Journal of Nutrition, 110, 375 – 383. https://doi.org/10.1017/s0007114512004953.
- Venegas, D., De La Fuente, M., Landskron, G., González, M., Quera, R., Dijkstra, G., Harmsen, H., Faber, K., & Hermoso, M. (2019). Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Frontiers in Immunology, 10. https://doi.org/10.3389/fimmu.2019.00277.
- Du, Y., He, C., An, Y., Huang, Y., Zhang, H., Fu, W., Wang, M., Shan, Z., Xie, J., Yang, Y., & Zhao, B. (2024). The Role of Short Chain Fatty Acids in Inflammation and Body Health. International journal of molecular sciences, 25(13), 7379. https://doi.org/10.3390/ijms25137379
- Jiao, A., Yu, B., He, J., Yu, J., Zheng, P., Luo, Y., Luo, J., Yan, H., Wang, Q., Wang, H., Mao, X., & Chen, D. (2021). Sodium acetate, propionate, and butyrate reduce fat accumulation in mice via modulating appetite and relevant genes.. Nutrition, 87-88, 111198 . https://doi.org/10.1016/j.nut.2021.111198.
- Zhang, Y., Li, X., Huang, G., Wang, H., Chen, H., Su, Y., Yu, K., & Zhu, W. (2022). Propionate stimulates the secretion of satiety hormones and reduces acute appetite in a cecal fistula pig model. Animal Nutrition, 10, 390 – 398. https://doi.org/10.1016/j.aninu.2022.06.003.
- Lu, Y., Fan, C., Li, P., Lu, Y., Chang, X., & Qi, K. (2016). Short Chain Fatty Acids Prevent High-fat-diet-induced Obesity in Mice by Regulating G Protein-coupled Receptors and Gut Microbiota. Scientific Reports, 6. https://doi.org/10.1038/srep37589.
- Lin, H., Frassetto, A., Kowalik, E., Nawrocki, A., Lu, M., Kosinski, J., Hubert, J., Szeto, D., Yao, X., Forrest, G., & Marsh, D. (2012). Butyrate and Propionate Protect against Diet-Induced Obesity and Regulate Gut Hormones via Free Fatty Acid Receptor 3-Independent Mechanisms. PLoS ONE, 7. https://doi.org/10.1371/journal.pone.0035240.
- Blaak, E., Canfora, E., Theis, S., Frost, G., Groen, A., Mithieux, G., Nauta, A., Scott, K., Stahl, B., Van Harsselaar, J., Van Tol, R., Vaughan, E., & Verbeke, K. (2020). Short chain fatty acids in human gut and metabolic health.. Beneficial microbes, 1-46 . https://doi.org/10.3920/bm2020.0057.
- Darzi, J., Frost, G. S., & Robertson, M. D. (2011). Do SCFA have a role in appetite regulation?. The Proceedings of the Nutrition Society, 70(1), 119–128. https://doi.org/10.1017/S0029665110004039
- Chambers, E. S., Viardot, A., Psichas, A., Morrison, D. J., Murphy, K. G., Zac-Varghese, S. E., MacDougall, K., Preston, T., Tedford, C., Finlayson, G. S., Blundell, J. E., Bell, J. D., Thomas, E. L., Mt-Isa, S., Ashby, D., Gibson, G. R., Kolida, S., Dhillo, W. S., Bloom, S. R., Morley, W., … Frost, G. (2015). Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut, 64(11), 1744–1754. https://doi.org/10.1136/gutjnl-2014-307913
- Testa, R., Vitale, M., Giosuè, A., Salamone, D., Quaglia, C., Rivellese, A. A., Bozzetto, L., & Costabile, G. (2026). Targeting weight loss and blood glucose control with oral sodium butyrate in overweight/obese adults with and without type 2 diabetes: A proof-of-concept randomized controlled trial. Clinical nutrition (Edinburgh, Scotland), 60, 106624. https://doi.org/10.1016/j.clnu.2026.106624
- Huwiler, V. V., Schönenberger, K. A., Segesser von Brunegg, A., Reber, E., Mühlebach, S., Stanga, Z., & Balmer, M. L. (2022). Prolonged Isolated Soluble Dietary Fibre Supplementation in Overweight and Obese Patients: A Systematic Review with Meta-Analysis of Randomised Controlled Trials. Nutrients, 14(13), 2627. https://doi.org/10.3390/nu14132627
- Thompson, S. V., Hannon, B. A., An, R., & Holscher, H. D. (2017). Effects of isolated soluble fiber supplementation on body weight, glycemia, and insulinemia in adults with overweight and obesity: a systematic review and meta-analysis of randomized controlled trials. The American journal of clinical nutrition, 106(6), 1514–1528. https://doi.org/10.3945/ajcn.117.163246
- Liu, L., Wang, H., Chen, X., Zhang, Y., Zhang, H., & Xie, P. (2023). Gut microbiota and its metabolites in depression: from pathogenesis to treatment. eBioMedicine, 90. https://doi.org/10.1016/j.ebiom.2023.104527.
- Barandouzi, Z., Starkweather, A., Henderson, W., Gyamfi, A., & Cong, X. (2020). Altered Composition of Gut Microbiota in Depression: A Systematic Review. Frontiers in Psychiatry, 11. https://doi.org/10.3389/fpsyt.2020.00541.
- Osimo, E., Baxter, L., Lewis, G., Jones, P., & Khandaker, G. (2019). Prevalence of low-grade inflammation in depression: a systematic review and meta-analysis of CRP levels. Psychological Medicine, 49, 1958 – 1970. https://doi.org/10.1017/s0033291719001454.
- Hao, W., , Q., Wang, L., Yuan, N., Gan, H., He, L., Li, X., Huang, J., & Chen, J. (2024). Gut dysbiosis induces the development of depression-like behavior through abnormal synapse pruning in microglia-mediated by complement C3. Microbiome, 12. https://doi.org/10.1186/s40168-024-01756-6.
- Privitera, G., Misenheimer, M., & Doraiswamy, P. (2013). From weight loss to weight gain: appetite changes in major depressive disorder as a mirror into brain-environment interactions. Frontiers in Psychology, 4. https://doi.org/10.3389/fpsyg.2013.00873.
- Mills, J., Larkin, T., Deng, C., & Thomas, S. (2019). Weight gain in Major Depressive Disorder: Linking appetite and disordered eating to leptin and ghrelin.. Psychiatry research. https://doi.org/10.1016/j.psychres.2019.03.017.
- Carlsen, H., & Pajari, A. (2023). Dietary fiber – a scoping review for Nordic Nutrition Recommendations 2023. Food & Nutrition Research, 67. https://doi.org/10.29219/fnr.v67.9979.
- Rondinella, D., Raoul, P., Valeriani, E., Venturini, I., Cintoni, M., Severino, A., Galli, F., Mora, V., Mele, M., Cammarota, G., Gasbarrini, A., Rinninella, E., & Ianiro, G. (2025). The Detrimental Impact of Ultra-Processed Foods on the Human Gut Microbiome and Gut Barrier. Nutrients, 17. https://doi.org/10.3390/nu17050859.
- Whelan, K., Bancil, A., Lindsay, J., & Chassaing, B. (2024). Ultra-processed foods and food additives in gut health and disease. Nature Reviews Gastroenterology & Hepatology, 21, 406 – 427. https://doi.org/10.1038/s41575-024-00893-5.
- Varghese, S., Rao, S., Khattak, A., Zamir, F., & Chaari, A. (2024). Physical Exercise and the Gut Microbiome: A Bidirectional Relationship Influencing Health and Performance. Nutrients, 16. https://doi.org/10.3390/nu16213663.
- Clauss, M., Gérard, P., Mosca, A., & Leclerc, M. (2021). Interplay Between Exercise and Gut Microbiome in the Context of Human Health and Performance. Frontiers in Nutrition, 8. https://doi.org/10.3389/fnut.2021.637010.
- Bonomini-Gnutzmann, R., Plaza-Díaz, J., Jorquera-Aguilera, C., Rodríguez-Rodríguez, A., & Rodríguez-Rodríguez, F. (2022). Effect of Intensity and Duration of Exercise on Gut Microbiota in Humans: A Systematic Review. International Journal of Environmental Research and Public Health, 19. https://doi.org/10.3390/ijerph19159518.
- , L., Yan, Y., Webb, R., Li, Y., Mehrabani, S., Xin, B., Sun, X., Wang, Y., & Mazidi, M. (2023). Psychological Stress and Gut Microbiota Composition: A Systematic Review of Human Studies. Neuropsychobiology, 82, 247 – 262. https://doi.org/10.1159/000533131.
- (2022). Stressful events induce long-term gut microbiota dysbiosis and associated post-traumatic stress symptoms in healthcare workers fighting against COVID-19. Journal of Affective Disorders, 303, 187 – 195. https://doi.org/10.1016/j.jad.2022.02.024.
- Allen, J., Mackos, A., Jaggers, R., Brewster, P., Webb, M., Lin, C., Ladaika, C., Davies, R., White, P., Loman, B., & Bailey, M. (2021). Psychological stress disrupts intestinal epithelial cell function and mucosal integrity through microbe and host-directed processes. Gut Microbes, 14. https://doi.org/10.1080/19490976.2022.2035661.
- Morys, J., Małecki, A., & Nowacka-Chmielewska, M. (2024). Stress and the gut-brain axis: an inflammatory perspective. Frontiers in Molecular Neuroscience, 17. https://doi.org/10.3389/fnmol.2024.1415567.
- Zhou, X., Ganz, A., Lu, X., Li, Y., & Snyder, M. (2023). Association between the dynamics of the gut microbiota and responsiveness to mental health therapy. The Journal of Immunology. https://doi.org/10.4049/jimmunol.210.supp.241.16.
- Sun, Y., Ju, P., Xue, T., Ali, U., Cui, D., & Chen, J. (2023). Alteration of faecal microbiota balance related to long-term deep meditation. General Psychiatry, 36. https://doi.org/10.1136/gpsych-2022-100893.
- Matenchuk, B., Mandhane, P., & Kozyrskyj, A. (2020). Sleep, circadian rhythm, and gut microbiota.. Sleep medicine reviews, 53, 101340 . https://doi.org/10.1016/j.smrv.2020.101340.
- Shan, W., Zang, W., & Zuo, Z. (2025). Sleep deprivation disrupts diurnal rhythmicity of gut microbiota and blood inflammatory cytokines in mice. PLOS One, 20. https://doi.org/10.1371/journal.pone.0335754.
- Wang, Z., Chen, W., Li, S., He, Z., Zhu, W., Ji, Y., Wang, Z., Zhu, X., Yuan, K., Bao, Y., Shi, L., Meng, S., Xue, Y., Xie, W., Shi, J., Yan, W., Wei, H., Lu, L., & Han, Y. (2021). Gut microbiota modulates the inflammatory response and cognitive impairment induced by sleep deprivation. Molecular Psychiatry, 26, 6277 – 6292. https://doi.org/10.1038/s41380-021-01113-1.
- Hong, S., Lee, D., Yoon, D., Yoo, S., & Kim, J. (2025). The Effect of Sleep Disruption on Cardiometabolic Health. Life, 15. https://doi.org/10.3390/life15010060.
- Savin, Z., Kivity, S., Yonath, H., & Yehuda, S. (2018). Smoking and the intestinal microbiome. Archives of Microbiology, 200, 677 – 684. https://doi.org/10.1007/s00203-018-1506-2.
- Antinozzi, M., Giffi, M., Sini, N., Gallé, F., Valeriani, F., De Vito, C., Liguori, G., Spica, R., & Cattaruzza, M. (2022). Cigarette Smoking and Human Gut Microbiota in Healthy Adults: A Systematic Review. Biomedicines, 10. https://doi.org/10.3390/biomedicines10020510.
- Koponen, K., McDonald, D., Jousilahti, P., Méric, G., Inouye, M., Lahti, L., Niiranen, T., Männistö, S., Havulinna, A., Knight, R., & Salomaa, V. (2025). Associations of alcohol with the human gut microbiome and prospective health outcomes in the FINRISK 2002 cohort. European Journal of Nutrition, 64. https://doi.org/10.1007/s00394-025-03668-z.
- Kosnicki, K., Penprase, J., Cintora, P., Torres, P., Harris, G., Brasser, S., & Kelley, S. (2019). Effects of moderate, voluntary ethanol consumption on the rat and human gut microbiome. Addiction Biology, 24, 617 – 630. https://doi.org/10.1111/adb.12626.
- Martínez-Montoro, J., Quesada-Molina, M., Gutiérrez‐Repiso, C., Ruiz-Limón, P., Subiri-Verdugo, A., Tinahones, F., & Moreno-Indias, I. (2022). Effect of Moderate Consumption of Different Phenolic-Content Beers on the Human Gut Microbiota Composition: A Randomized Crossover Trial. Antioxidants, 11. https://doi.org/10.3390/antiox11040696.
- González-Zancada, N., Redondo-Useros, N., Díaz, L., Gómez-Martínez, S., Marcos, A., & Nova, E. (2020). Association of Moderate Beer Consumption with the Gut Microbiota and SCFA of Healthy Adults. Molecules, 25. https://doi.org/10.3390/molecules25204772.
- Liu, Z., Shi, J., Wang, L., Dong, J., Yu, J., & Chen, M. (2024). Association of Moderate Beer Consumption with the Gut Microbiota. Food Science and Human Wellness. https://doi.org/10.26599/fshw.2023.9250004.
- Zimmermann, P., & Curtis, N. (2019). The effect of antibiotics on the composition of the intestinal microbiota.. The Journal of infection. https://doi.org/10.1016/j.jinf.2019.10.008.
- Imhann, F., Bonder, M., Vila, V., Fu, J., Mujagic, Z., Vork, L., Tigchelaar, E., Jankipersadsing, S., Cenit, M., Harmsen, H., Dijkstra, G., Franke, L., Xavier, R., Jonkers, D., Wijmenga, C., Weersma, R., & Zhernakova, A. (2015). Proton pump inhibitors affect the gut microbiome. Gut, 65, 740 – 748. https://doi.org/10.1136/gutjnl-2015-310376.